
Cane toads and advancing range edges

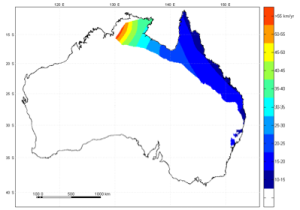
In northern Australia, the cane toad front pushes westward at around 55km per year; more than five times faster than it used to advance when toads were first introduced. This staggering rate of population advance means that individual toads in northern Australia often move more than 55km in a year. There is no other amphibian in the world that comes close to these rates of movement. Why do toads do this? Apart from increased mortality associated with increased activity, toads on the invasion front in northern Australia also suffer a heightened prevalence of spinal arthritis as a consequence of their excessive nightly wanderings. Additionally, it makes very little sense for an amphibian (prone to dessication) to wander off into unknown territory with no guarantee of finding the next waterhole. So why do they do it?
Dispersal Evolution and the Olympic Village Effect
The answer is simple: they can't help themselves. Via a process of assortative mating, cane toads on the leading edge of the invasion front have been continually selected for increased dispersal ability. How so? If we imagine a population where all the individuals disperse, then reproduce, and then die, and that this population is placed into a large area of suitable vacant habitat, the population will advance its range as individuals disperse. Importantly, the process of dispersal effectively sorts individuals through space by dispersal ability. That is, individuals on the edge of the expanding population front are at that edge simply because they dispersed further than other individuals in the population: the process of dispersal has spatially assorted the best dispersers in the population and placed them on the expanding front. Now, because all the best dispersers are in the same place at the same time, they will tend to breed with each other (the Olympic Village effect). Thus, if any component of dispersal ability is heritable in this hypothetical species, the offspring of the individuals on the front will tend to have higher dispersal ability than the offspring of individuals from the core of the population. If we imagine this process occurring every generation as the population expands, the process of spatial assortment for dispersal ability behaves like a runaway evolutionary process, continually selecting for increased dispersal on the advancing front.
Evolution of reproductive rate during range advance
Interacting with this process of spatial assortment are density effects. That is, individuals near the advancing front may benefit from a low density environment (i.e., fewer conspecifics in their vicinity) and, as a consequence, leave more offspring. In fact, the range edge population will tend to grow exponentially, whereas further back in the range, the population will likely be density regulated. This has subtle but important implications for the evolution of reproductive rate on expanding range edges. In exponentially growing populations, any individual with a slight advantage in reproductive rate (offspring per unit time) will soon have its genes numerically dominate the population. This is classical r-selection. Further back in the range, we might expect the population to be under less intense r-selection. Thus, the process of range advance not only selects for increased dispersal, but also for increased reproductive rates.
Importantly, dispersal and reproductive rates are the two things that determine the rate of range expansion. Thus, toads have shown us that, for any species, range advance should accelerate over time because of directional and predictable evolutionary shifts. Good news for species trying to track climate change; bad news for people trying to stop the spread of invasives!